Рецессивный
аллель этого локуса носит название "неагути" (обозначается
а) и
обеспечивает однотонный окрас волосков – именно волосков, но не
обязательно кошки в целом. То есть кошка с генотипом аа может быть
черной, шоколадной, кремовой или даже иметь окрас блю-пойнт - в
зависимости от того, какие аллели будут входить в другие локусы
генов, ответственных за окрас. Естественно, что у кошек с окрасом
агути цвет полос на волосе тоже зависит от аллельного состояния тех
же генов - могут чередоваться полосы черные с желтыми, голубые с
желтоватыми, кирпичные со светло-рыжими и так далее. Продолжение статьи:
Каким
же образом может работать агути-ген? Вероятно, чередование отложения
пигментов происходит при росте волоса по принципу "встроенных"
биологических часов, с периодическим включением и выключением генной
активности. Можно отметить еще и совершенно различный характер в
ширине и количестве полосок тикинга у разных кошек. У других
млекопитающих, имеющих подобные окрасы, например мышей и собак,
агути считается сложным, комплексным геном и представлен не двумя
аллелями, а обширными аллельными сериями.
Если кошки с
генотипом неагути обычно имеют однотонную окраску (не считая
акромеланистических и дымчатых вариаций), то окрасы агути, как
правило, сочетаются с тем или иным рисунком на теле кошки. За
наличие и тип такого рисунка отвечают аллели серии тэбби (Т -
Tabby).
Изредка, правда, в естественных популяциях встречаются кошки с
ярко выраженным тикингом, но практически без рисунка. Тот же окрас
характеризует и абиссинскую породу кошек.
Доминантный,
аллель, отвечающий за формирование этого окраса, называемого
абиссинским или тикированным тэбби, обозначается как Та. Впрочем, гетерозиготы по этому гену, а изредка и гомозиготы -ТаТа
имеют
остаточные элементы рисунка: кольца "ожерелья"
на груди, слабые
полосы на ногах и отметину в форме буквы "М" на лбу.
У
кошек России наиболее распространен тигровый рисунок (или
макрель-тэбби) - то есть вертикальные полосы на туловище в
сочетании с общими для всех рисунчатых окрасов элементами: отметина
"М" на лбу, кольца на груди, завитки на щеках, два ряда двойных
пятнышек вдоль живота и полосы на хвосте и ногах. Такой рисунок
определяется аллелем Т, рецессивным по отношению к Та. А вот
довольно редкий у нас, но обычный для Великобритании мраморный окрас
кошек ( включает "бабочку" на плечах, две полосы вдоль спины и
разводы на боках) характерен для гомозигот по наиболее рецессивному
аллелю серии тэбби -
tbtb.
Самым
загадочным рисунком остается пятнистый (spotted)
состоящий в наиболее законченном выражении из ровных, одинаковых по
размеру круглых или овальных пятнышек на боках (естественно, в
сочетании с общими для всех рисунков элементами). При скрещивании
между собой пятнистые кошки всегда дают котят только с таким же
рисунком и никогда - тигровых. Неизвестны и случаи появления
пятнистых котят у пар мраморного окраса. Казалось бы, пятнистый
рисунок должен определяться аллелем той же серии тэбби, рецессивно
наследуемым по отношению к Т и доминантным - по отношению к
tb,
и обозначаться как-то вроде
tsp.
Между тигровым и пятнистым окрасам наблюдаются все переходные
степени - от разорванных в нескольких местах полос до почти круглых
пятен. Известны и такие особи, у которых 2-3 полосы на передней
части тела сочетаются с пятнами на задней. Это могло бы быть
объяснено промежуточным проявлением аллелей Т и
tsp.
Интересно, что на сотни кошек с промежуточными тигрово-пятнистыми
окрасами известны буквально единицы с разорванным мраморным
рисунком! Возможным объяснением этого явления может быть то, что
тэбби представляет собой сложный, протяженный локус, внутри которого
возможен обмен участками - генетическим материалом (внутригенная
рекомбинация). Другая гипотеза о происхождении пятнистых окрасов
предполагает, что разрывы в тигровом окрасе и - как крайняя форма
этого явления – пятнистость, вызваны действием генов другого,
независимого от тэбби локуса. Но последнее предположение также не
объясняет количественной разницы между разрывами полос тигрового и
мраморного рисунков.
Механизм
развития рисунка, скорее всего, тоже связан с внутренними
биологическими часами развития организма. Даже у маленьких белых
котят можно заметить такой "рисунок", образованный полосками волос
разной длины и текстуры: более длинные и грубые чередуются с более
тонкими и короткими.
Вероятно,
разное окрашивание волосков рисунка (они более темные, прокрашены
целиком или с широкими полосами темного пигмента) и волосков фона (с
более широкими полосами феомеланина) связано с разными сроками
созревания волосяных фолликулов, разной скоростью роста волос, а
значит, и с разными сроками включения-выключения генов,
ответственных за синтез пигментов.
Тем
не менее существуют и такие мутации, которые могут не только
изменить всю картину чередования черно-желтых полос, но и полностью
"смыть" рисунок с тела кошки, несмотря на ее генотип агути.
Наследование таких окрасов - одна из интереснейших проблем генетики
кошек. Долгие годы считалось, что появление их обусловлено действием
гена-ингибитора меланина -
I.
Рецессивный аллель этого локуса-
i
- никакого внешне заметного влияния на синтез пигмента не оказывает,
а доминантный аллель того же гена останавливает синтез меланина
таким образом, что у остевых волос окрашивается только верхняя
часть, а основания ости и подшерсток кошки вообще остаются целыми.
Однако
быстро выяснилось, что называть аллель I доминантным не совсем
правильно. Дело в том, что его экспрессивность изменяется в очень
широких пределах. Предполагалось, что деятельность гена-ингибитора
лежит в основе нескольких групп окрасов. На генетическом фоне
неагути — аа — остевые волоски под действием этого гена
непрокрашиваются почти на половину длины, а подшерсток остается
полностью белым. Такой окрас кошек носит название дымчатого. Но
нередко встречаются дымчатые окрасы с плохо выбеленным, сероватым
подшерстком.
У серебристых
тэбби, окрасах, которые развиваются под действием гена-ингибитора на
основе генотипа -А-, волоски в рисунке часто прокрашены почти до
основания, тогда как у остевой шерсти фона цветными остаются только
кончики. Причем очень часто и у дымчатых кошек просвечивает теневой
рисунок, и волоски в нем более темные. Особенно ярко это явление
выражено у котят, и маленьких "дымов" путают с серебристыми тэбби.
Крайняя
степень активности гена-ингибитора - это так называемые затушеванные
и затененные окрасы (шиншиллы). Эти окрасы также развиваются на
генетическом фоне агути. У первых кончик волоса прокрашен примерно
на 1/3 длины, а у вторых - всего на 1/8, без каких бы то ни было
полосок. Такое распределение цвета по волосу носит название типинга.
Естественно, цвет кончиков волос зависит от того, какие аллели
содержатся в локусах
B,
D
и
O.
К названиям окрасов
затененных и
затушеванных кошек с красными или кремовыми кончиками волос
добавляется слово "камео".
Описанные
вариации проявления ингибитора меланина дают основание предполагать
куда более сложную картину взаимодействия генов, чем влияние только
одного аллеля I. Тем более что к серебристой группе окрасов,
связанных с процессами частичного нарушения синтеза пигментов,
добавились и золотистые.
Первый
и основной признак золотистого окраса: от 1/2 (золотые тэбби) до 2/3
(золотые затушеванные) или 7/8 (шиншиллы) часть каждого остевого и
покровного волоска окрашены в светло- или ярко-абрикосовый, теплый
тон. Оттенки этого тона на разных участках тела кошки могут
различаться, но ни в коем случае не должны переходить в тусклые,
сероватые цвета. Наиболее частым (не сказать, чтоб приятным)
дополнением к окрасу золотых тэбби и золотых затушеванных кошек
служат остаточные полосы тикинга на прокрашенной в темной части
остевых волосков, что или "смазывает" рисунок (у тэбби), или придает
неряшливый вид окрасу (у затушеванных). Этот недостаток настолько
распространен, что рассматривается чуть ли не как норма.
Крайне
часто встречаются цветовые вариации кошек, промежуточные между
золотыми и обычными черными тэбби: остевые волосы таких животных
прокрашены в "золото", а вот подшерсток - серый. Обычно и глаза этих
особей не достигают изумрудно-зеленого цвета, характерного для
золотых окрасов.
Среди золотых
кошек с рисунком (тэбби) наблюдается и еще одна вариация золотого
окраса - когда и подшерсток золотой, и фон ости хорошо высветлен, а
вот покровные волоски в рисунке затемнены почти до корней. У кошек
такого типа не отмечается полосок тикинга в рисунке, и собственно
"золото" интенсивного, почти медного цвета, что представляет собой
явно положительное качество. К сожалению, выборка кошек этого
последнего типа крайне мала. Итак, среди золотых окрасов можно
выделить как минимум три различных типа, а также все варианты
переходов между ними.
Впервые
помет золотистых кошек-шиншилл был получен от серебристых
шиншилл-родителей. Поэтому первоначально считали что золотистый
окрас определяется наличием того же полудоминантного гена-ингибитора
(генетический символ I), который обеспечивает серебристые окрасы
шиншилл, затушеванных, тэбби и дымчатых кошек.
Однако
работой одного лишь гена, хотя бы и полудоминантного, всех вариаций
окрасов, получаемых в серебристо-золотой гамме, объяснить не
удавалось. Поэтому генетиками было выдвинуто предположение о генах
руфизма — то есть группе генов, обеспечивающий дополнительный синтез
желтого пигмента - феомеланина. Но и это слишком расплывчатое
предположение не было признано удовлетворительным.
Пусть
и не самых совершенных проявлениях, но золотистый окрас довольно
распространен в кошачьих популяциях. Поиск генов, отвечающих за
столь соблазнительный окрас, был продолжен. Исследователи прежде
всего обратили внимание на так называемые "вавиловские ряды", то
есть сходство в мутациях окрасов у различных групп животных:
например, и сиамские коты, и гималайские кролики, и
акромеланисты-мыши - все они имеют одинаково определяемый
генетически окрас.
Согласно
этому закону параллелизма, на роль кандидатов в гены золотого окраса
был выдвинут доминантный ген "широкой полосы" —
Wb
обнаруженный у некоторых грызунов. Под действием этого гена
формируется широкая желтая полоса в основании волоса, и животное
приобретает золотой окрас. В случае действия нормального аллеля гена
wb
получается обычный черный тэбби, если же к этому генетическому фону
добавить ген-ингибитор, то формируется серебристый тэбби.
Когда
же аллели I и
Wb
сосредоточены в одном организме, образуются серебристые шиншиллы или
затушеванные. Другая гипотеза, также основанная, на параллелизме
окрасов, заключается в наличии у кошек гена "золотого агути"
(генетический символ Ау), характерного для собак и мышей. У
большинства хорошо изученных генетически млекопитающих
агути-комплекс представлен не только двумя аллелями, то есть
вариантами гена, известными у кошек (А - агути и а - неагути), а
целой серией аллелей. Так называемый "соболиный" окрас собак,
например, связан именно с действием аллеля "золотого агути" и
заключается в желтом прокрашивании волос (за исключением их темных
кончиков). Если исходить из предположения о наличии такого же гена у
кошек, то дальнейшие рассуждения о формировании серебристо-золотой
гаммы окрасов окажутся сходными с изложенными выше, с той
разницей, что место гипотетического рецессивного
wb
будет занимать привычный агути-фактор Ау.
В
настоящее время наиболее распространены бигенные теории золотых и
серебристых окрасов, то есть базирующиеся на двух отдельных локусах
(или генетических комплексах).
Чтобы
ознакомиться с одной из последних теорий наследования золотых и
серебристых окрасов, основанной на взаимодействии двух независимых
генов на фоне агути или неагути-мутации, следует вспомнить некоторые
особенности разведения не только этих, но и дымчатых окрасов:
-
при
скрещивании золотистых тэбби или затушеванных кошек между собой не
появляется серебристого потомства, тогда как появление золотистых
затушеванных при скрещивании серебристых шиншилл — случай довольно
обычный; -
серебристые кошки с рисунком при скрещивании могут дать золотистое
потомство только в том случае, если серебро родителей недостаточно
качественное - отмечается желтый тикинг в рисунке, желтые надцветы
на мордочке и прочие недостатки окраса; -
при
инбредном разведении (родственных скрещиваниях кошек с ярко
выраженным золотистым окрасом рождаются золотистые потомки (иногда выщепляются осветленные);
-
при
неродственных скрещиваниях золотистых кошек, а также при
скрещивании их с серебристыми среди золотистых потомков нередко
встречаются котята с серым и бурым подшерстком, а среди серебристых
- с желтоватым тикингом по волоскам и желтыми надцветами на мордочке
и лапах; -
при
скрещивании золотистых кошек с черными тэбби все потомство или по
крайней мере половина его - обычные черные тэбби, но обнаруживаются
и потомки промежуточных окрасов, причем у таких особей подшерсток
обычно серый, а "золото" заметно только в остевых волосках;
-
при
неродственных скрещиваниях дымчатых кошек между собой или с
однотонными часто появляются потомки со светло-серым "холодным"
подшерстком; -
с
другой стороны, среди однотонных кошек нередко встречаются особи с
теплым рыжеватым надцветом на шерсти и тоном подшерстка.
Остается
предположить, что гены, отвечающие за серебристый окрас (ингибиторы
меланина, и прежде всего его желтой модификации - феомеланина)
действуют независимо от генов золотого окраса - ингибиторов
эумеланина, черного пигмента (на то, что ген золотого окраса - также
ингибитор пигмента, указывает и корреляция окраса с зеленым -
недопрокрашенным - цветом глаз). В одной из последних работ эти гены
были соответственно названы
Bleacher
и
Eraser
(название и генетические символы неофициальные). Каждый из этих
генов должен быть представлен как минимум двумя аллелями,
действующими, естественно, на агути или неагути-фоне.
Условно
принято, что эти гены обладают одинаковой генетической активностью.
В реальности, конечно, соотношения доминантности - рецессивности не
соблюдаются так уж строго и проявление генов варьирует в довольно
широких пределах, доказательством чего могут служить нередко
отмечавшиеся промежуточные цветовые формы. Взаимодействие этих
генов-ингибиторов проще проиллюстрировать таблицей (табл. 3).Смотрите ниже 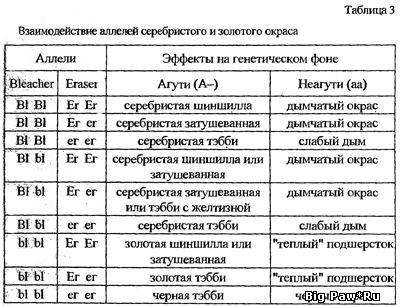
Кроме
того, известно, что степень проявления гена часто зависит от его
дозы, то есть количества копий.
То есть серебристая гомозиготная
кошка будет обладать более ярко выраженным "серебром", чем
гетерозиготная. При этом следует учесть способность генов
удваиваться, увеличивать свою копийность в результате мутаций.
Естественно, желательные комбинации окраса немедленно закрепляются
заводчиками, и таким образом количество копий гена в популяции или
питомнике возрастает. Что же касается генов-модификаторов руфизма,
то их роль теперь определяется как факторов, модифицирующих
интенсивность желтого пигмента - от бледно-золотистого до
ярко-медного. Вероятно, их действие связано или с интенсивностью
синтеза феомеланина, или со степенью его концентрации в волосяных
фолликулах.
Самостоятельного
генетического символа эти гены не имеют и существуют, так сказать,
"на птичьих правах".
Приведенная выше
схема не может, конечно, ответить на все вопросы формирования
золотых, серебристых и дымчатых окрасов - почему, например,
существует столь явная корреляция между тикингом ости и золотым
окрасом основания волоса? Возможно, что составляющие агути-комплекса
играют для этих окрасов не только статическую роль генетического
фона, но и непосредственно вовлечены в формирование золотого (то
есть лишенного эумеланина) тона шерсти. То есть, помимо известных
нам двух аллельных состояний агути-гена (А и а), существуют и другие
представители этой генетической серии, подобно описанному выше аллелю "желтого агути". Проявление генов - ингибиторов эу- и
феомеланина при взаимодействии с иными агути-аллелями и может давать
те самые, пока не объясненные эффекты золотых окрасов.
Составление
и чтение генетических формул окраса
После
знакомства с генетикой пигментогенеза составление и чтение
генетических формул окраса не составит большого труда. Часть
генотипа кошки записана, так сказать, на ней.
Возьмем
для примера голубокремовую кошку. Ее окрас не белый - следовательно,
она гомозиготна по рецессивному аллелю гена
W
-
ww,
белые пятна отсутствуют -
ss,
агути-фактор не выражен – аа.
Далее, нам
известно, что голубой окрас — это осветленный черный. Осветленный
записывается просто — dd.
Относительно
черного можно отметить только один доминантный аллель — В-.
Состояние же его гомолога по внешнему виду кошки определить
невозможно, она может быть и гомо-, и гетерозиготной. То же касается
гена альбинизма — мы видим полное окрашивание — (С-), но ничто не
мешает этой кошке в своем генотипе нести аллель с или cs. Поскольку
кошка черепаховая, ясно, что она гетерозиготна по гену красного
окраса — Оо. Отсутствие признаков "дыма" можно записать как
ii
или как
erer. А
вот определить на черепаховой кошке, "теплый" ли у нее тон
подшерстка, практически нереально.
Точно
так же по ее внешнему виду нельзя судить о том, какой тип рисунка
закодирован в ее генотипе (но это не означает, что у кошки нет
никаких аллелей в локусе Т или самого этого локуса!). Итак, судя по
"внешнему виду кошки, ее генотип должен быть записан как: ааВ-С-ddOosswwii(или
erer).
Часть
данных о генотипе животного мы можем вывести из его родословной,
точнее, установить по внешнему облику родителей. Если мать
описываемой кошки была шоколадно-мраморная, а отец кремовый
мраморный, то аллельное состояние локусов В и Т можно определить как
гетерозиготное - Bb и гомозиготное - tbtb. Но если кто-то из
родителей имел тигровый рисунок, то мы не можем записать состояние
гена как
Ttb,
так как не знаем, была ли мать гомозиготной по доминантному аллелю
Т
и передала ли своей дочери именно этот аллель, а не его рецессивный
гомолог. Конечно, если отец или мать кошки имеют белый окрас, то
никакой информации о состоянии интересующих нас генов получить
невозможно. А вот если при наличии шоколадно-мраморной матери отец
кошки в родословной записан как лиловый или черный, то мы, скорее
всего, имеем дело с подделкой: от таких родителей кошка никак не
могла унаследовать доминантный ген О.
Генетические
формулы можно использовать для расчета вероятности получения тех или
окрасов при вязках. В последние годы появились целые книги таблиц,
где приводятся возможные окрасы потомства при различных цветовых
вариантах вязок. Однако любой мало-мальски генетически грамотный
заводчик легко проведет эти расчеты и без помощи справочной таблицы.
Во-первых, следует определить генотипы родителей (так, как это было
сделано выше). Допустим, вышеописанную голубокремовую самку мы хотим
повязать шоколадным котом, происходящим от лилового отца и черной
матери. Генотип кота:
aabbC-DdoY
(то есть Y-хромосома)
sswwii.
При составлении таблицы расчета (а за ее основу взята хорошо нам
знакомая решетка Пеннета) можно пренебречь теми генами, аллельное
состояние которых одинаково у обоих родителей. У потомков оно,
конечно, не изменится. Что же касается тех генов, состояние которых
нам неизвестно - С, Т и Вl -то двумя последними в этом конкретном
случае также можно пренебречь, поскольку от двух особей без
агути-фактора ни тэбби, ни шиншилл получить невозможно. Ген С мы
тоже пока не будем включать в таблицу, однако вернемся к его
рассмотрению позже. Так же, как и в случае с решеткой Пеннета, по
горизонтали размещаем варианты генетического набора, образующиеся в
половых клетках одного из родителей, по вертикали - другого. На
пересечении записываем генотип котенка. По генетической формуле уже
легко прочитать окрас (табл. 4). Смотрите ниже 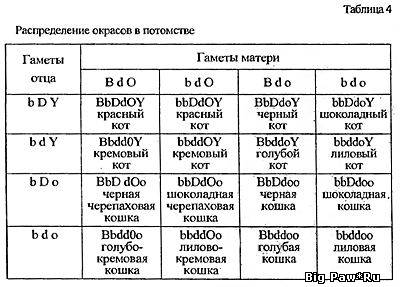
Генотипы
используемой в скрещивании пары можно уточнить не только исходя из
внешности их родителей, но и по потомству. Если от интересующих нас
особей родился хотя бы один котенок-колорпойнт, то можно утверждать,
что оба животных гетерозиготны по генам локуса С:
cscs.
Соотношение этих акромеланистических котят с полностью окрашенными
составляет примерно 1:3, а вероятное распределение окрасов среди них
в точности повторяет уже просчитанное нами скрещивание (табл. 5).Смотрите ниже 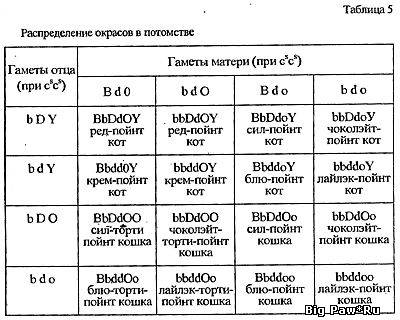
Разумеется,
при таких расчетах надо помнить, что количественное соотношение
окрасов котят - величина вероятностная, оно может выполняться только
при большой выборке, которую нереально получить от одной пары
животных.
Источник: http://www.vandvis.ru/index.htm |
